風水地理-凹岸凸岸
http://blog.xuite.net/yuchen99/twblog/157749708-%E5%85%AB%E5%85%AB%E6%B0%B4%E7%81%BD%EF%BC%8D%E9%87%91%E5%B8%A5%E9%A3%AF%E5%BA%97%E5%80%92%E5%A1%8C
島嶼生物地理學與應用
http://geog-education.blogspot.tw/2010/11/blog-post_13.html#more/游牧笛
島嶼生物地理學(Island Biogeography)顧名思義,就是一門專門研究被海環繞的島嶼,其生物地理分佈特性的學門。島嶼生物地理學在近年來受到了廣大的關注,以鳥類來說,雖然全世界的島嶼上鳥類種數、亞種數僅佔全世界的20%,但是西元1600年以來,滅絕的171個鳥群(或亞種),卻有高達155個類群(約90%)是僅分布於島嶼的種類,島嶼鳥類滅絕的速率遠超過大陸地區50倍,一定是有什麼機制而造成了這樣的結果。
在了解島嶼生物地理學之前,要先了解下列幾個重點:
1.島嶼的類型
島嶼大致上可以分成三類,第一類是大陸島,此類島嶼的地質構造與鄰近的大陸相近,是因為海平面上升,或是因為板塊張裂才與大陸分隔,例如東印度群島中的新幾內亞島、非洲外海的馬達加斯加島,還有我們居住的台灣島都屬於此類(紐西蘭島則屬於板塊張裂所形成的島嶼);第二類是板塊隱沒帶附近的島弧,例如阿留申群島等;第三類則是熱點島嶼,由地底的熱點(hot spot)所噴發出來的火山所形成,有許多位於大洋中央,例如夏威夷群島等大洋州中的「高島」。
此三種島嶼所擁有的生物相並不相同,大陸島的生物相與大陸本身相近,但又因為海水隔離而有一定程度的特化;島弧與熱點所生成的島嶼,則受到大洋生物相的影響較大。倘若大陸島因為某些原因而距離大陸本身越來越遠,則受到大洋生物相的影響就會逐漸加大;反之,若島弧或熱點島嶼因板塊漂移而移向大陸,那麼受到大陸生物相的影響也會逐漸變大。而當島弧、熱點島嶼與大陸或大陸島碰撞後,該區的生物相就會相當的複雜而多樣,這或許也可以說明台灣生物多樣性的來源。
2.生物如何到達島嶼
海洋,是動物在島嶼、大陸間遷徙最大的障礙,大部分的生物在海洋中遷徙靠的是所謂的「樂透途徑(sweepstakes route)」,意即強風吹送或者植物殘骸搭載,而隨機抵達島嶼。不同種類的動物在這樣的遷徙過程中,也有不同的極限,大部分的動物可能都無法撐過長時間的海洋輸送,但有部分生物卻可以長途跋涉。1995年時,有兩個颶風橫掃加勒比海,其後有一批木頭及被連根拔起的樹幹在安哥拉島(Anguilla)的海岸出現,裡頭有大約15隻綠鬣蜥,這些綠鬣蜥於兩年半後仍舊生存,且擁有可繁殖的個體。此前安哥拉島並無綠鬣蜥的紀錄,學者由颶風路徑推斷,可能是從250公里外的瓜德羅普(Guadeloupe)所送來,旅程可能達一個月之久。
植物在島嶼間的擴散則遠較動物容易,但儘管如此,不論動植物,島嶼的生物相仍強烈受到海洋隔閡的影響,短期而言,島嶼上生命的類別主要取決於動植物到達島嶼的速率,而這個速率則受到該島嶼與生物來源地的距離,以及生物來源地物種豐富度的影響。理論上,靠近生物種數豐富的大陸地區的島嶼,其生物種數也會較為豐富,而這個島嶼又成為下一個更深入海中的島嶼的生物來源地,而生物種數也逐漸遞減。
3.島嶼面積
島嶼的環境遠比大陸地區來的凶險,其主要原因可能是偶發性的災難,如毀滅性的風災、海嘯或火山爆發,也有長時距的影響,如氣候變遷等。在大陸上,某一區域特定種類的滅絕,可藉由其他地區同種類生物的遷入而獲得補償,但在島嶼上,由於整體環境相對封閉,一旦物種滅絕,很難再有其他族群遷入。
物種是否能順利在島嶼生存,是生物對於島嶼環境適應能力的唯一指標,然而,適應島嶼環境並不是一件簡單的事情。首先,如果從外地遷移來島嶼的族群數量太少,該族群所擁有能適應環境的基因數也就相對較少(奠基者效應:founder effect),而且小族群對非適應性的遺傳組成改變非常敏感,容易發生隨機性的滅絕(小族群基因滅絕:genetic drift)。島嶼物種難以生存,又可以從鳥類滅絕狀態看出,75%於島嶼滅絕的鳥種,是分布於小型島嶼的,因此「島嶼面積」對於物種滅絕速率,有絕對的影響。
綜合上述三點,我們可以簡單的描述幾個推論:
1.島嶼類型將會影響島嶼與大陸間的距離,以及島嶼面積的大小。
2.島嶼與生物來源地的距離,和物種遷入的速率約略呈反比。
3.島嶼面積大小,與物種的滅絕速率約略呈反比,島嶼越大,滅絕速率越慢。
有了上述的推論,島嶼生物地理學者便試圖整理出一套模式,藉以表現島嶼生物分布的特性。1967年美國生態學者R. McArthur和E. Wilson在《The Theory of Island Biogeography》一書中,利用量化理論來描述此現象。他們提出兩個結論:第一,移入和滅絕這兩個具有相關性的變動,最終將達到平衡;第二,島嶼面積與其擁有的物種數目有很強的相關性。
他們整理出來的結果,如下圖一般呈現:

他們以新形成、可供生物進駐的島嶼為例子作說明。在一開始,外來物種擴散繁殖的速率相當快速,因為擅於擴散的物種很快就會來到島上;隨著時間的演進,移入的生物越來越多是先前已經移入的種類,因此新種的增加速率會下降。此外,移入速率也受到島嶼位置的影響,較接近生物來源區的島嶼,生物擴散速度較快,而距離遠者擴散遷移進入速率較慢。
相反的,生物滅絕的速率一開始較低,因為一開始種類數不多,然而移入的物種越多,每種生物都有滅絕的可能性,因此滅絕速率會開始提高。此外,剛開始生物種類少,每種生物所能佔有的生態棲位(niche)也較為多樣,利於生存;隨著種類數越多,彼此競爭生態棲位的結果,將導致物種族群數目縮小,更增加了族群滅絕的危機。
假設島嶼上本來就有一種生物佔據A棲位,而又遷入了另一種同樣生存於A棲位的生物,兩種生物彼此的競爭,將可能導致其中一種物種完全滅絕於該島嶼,或者兩物種的族群數量都縮小而共有棲位(這會使得這兩種物種都更加特化,而生態棲位也更為狹窄,進而劃分兩種不同物種的棲位,但棲位縮小將使滅絕風險增加)。不論是上述哪種結果,都將造成滅絕速率的增加。
根據上述結論,R. McArthur和E. Wilson認為生物區系最後將達到平衡,遷入與滅絕速率大致相等,呈現的是一種動態平衡,彼此間以物種替換(species turnover)來維持。而此理論目前大致上被廣泛接受,儘管如此,也有不少學者提出異議,例如說蝸牛所需的生活範圍非常小,幾乎在任何島嶼都不會達到數量飽和,還有一些島嶼上的生物種數並找不到趨向於平衡的證據,但上述島嶼生物地理學的理論,提供了學者們比較他們自己研究的結果的平台,而跳脫出個案式的研究,此外,該理論也被援引到其他隔離環境,如地下洞穴、山峰頂端等。
近年來,島嶼生物地理學則提供了「保護區設立」一個方向,保護區的設立基本上是為了維持最大物種數量,在島嶼生物地理學理論中對於面積的描述,使得保護區、國家公園等的大小提供了一些想法,當然這還要配合保護區的保全對象特性。不消說,島嶼生態在保育上本來就有相當高的價值了。
臺灣植物垂直分布、北降和南降現象
http://geog-education.blogspot.tw/2010/12/blog-post.html#more/游牧笛
「北降現象」是一種植群的垂直分布因緯度升高,而逐漸降至低海拔的現象。其基本的外顯現象就是,在南邊(靠近赤道)必須生長在海拔較高處的植物,在北邊(距離赤道較遠)則可生長在比較低處;北降現象這個名詞的適用範圍僅限於北半球,在南半球則南北分布的狀態相反。
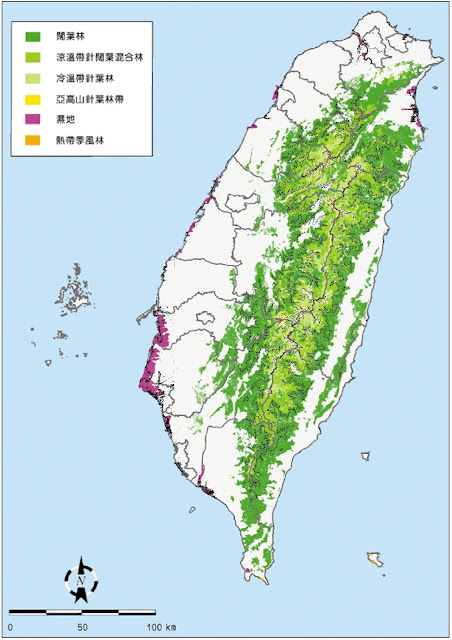
在細談北降現象之前,先來了解植物的垂直分布。在不考慮區域氣候差異的狀態下,垂直高度的變化將會導致植群分布的差異,在台灣(林務局),通常以嘉義到玉山一線的植群分布作為基準,分做熱帶林、副熱帶林、溫帶林、寒帶林以及高山草原五大部分;生態學者則以植群科別由低至高分為楠榕林帶、楠櫧林(樟殼林)、針闊葉混淆林(上、下部櫟林帶)、針葉林(鐵杉-雲山帶及冷杉帶),以及由草原(箭竹為主)組成的高山帶。當然還有很多不同的分帶法,因目的而異,不過都與「高度」有關。
會有上述的垂直分布,與熱能能量多寡有關,地表的熱能是地面長波輻射加熱為主(並不是陽光直射為主),因此海拔越高,長波輻射加熱能力越低,因此高海拔區域氣溫較低。同樣的,不同緯度帶的地表受熱程度也有相當差異,高緯度地區吸收的太陽短波輻射少,放出的地表長波輻射也就少,因此高緯度地區較為寒冷;正因為高緯度地區海拔1000公尺處可能與低緯度地區海拔2000公尺處的氣溫相同,因此在低緯度必須生長於2000公尺以上區域的植物,很有可能在高緯度地區海拔1000公尺處發現,此即為北降現象。
舉些簡單的例子,例如在嘉義-阿里山地區海拔2000公尺以上廣泛出現的紅檜林,在台灣北部的拉拉山、赫威、棲蘭山等紅檜林,海拔僅約1000多公尺。在台灣,除了受到緯度帶的影響外,冬季強大的東北季風造成的降溫,也增強了北降現象,如台灣中南部要在3000公尺以上才能廣泛看到的龍膽科植物,在台灣北部海拔約1000多公尺就可以見到。


華萊士線與其延伸
http://geog-education.blogspot.tw/2010/11/blog-post_12.html/游牧笛
生物的分布,一直是早期博物學家探索的主要課題之一,自從達爾文在加拉巴哥群島的發現震驚全世界之後,有更多的博物學家也投入了未知大陸的生物分佈研究。對地理學界而言,比較有名的大概就是洪堡德,這位醫生世家出身的學者,在南美洲進行了相當長一段的生物標本採集,也算是後來地理學界重視「實察」的鼻祖吧!
在地球的另一端,有些學者注意到,亞洲大陸東南側有連串的島嶼,好似把大陸陸塊延伸到另一頭的澳洲大陸,然而,澳洲大陸與亞洲大陸之間的生物相有極大的差異性,因此這些學者試圖在這些島中間,劃分出一條界線,以釐清哪些島的生物種類受到亞洲大陸影響較大?又有哪些島受到澳洲大陸的影響較大?
此時一位叫做華萊士(Alfred R. Wallace)的生物學家,在十八世紀時於印尼一帶做野外生物調查,分析陸生性動物在該地的動物相及群集分布。漸漸的在他蒐集的資料中,反映出生物在自然在地球上的分布。其中最明顯的動物相差異在峇里島(Bali)與龍目島(Lombok)之間。根據他的說法,「印尼峇里島的動物系以亞洲的要素的動物呈優勢,而龍目島以東以新幾內亞(New Guneia)要素的動物呈優勢(山崎柄根,2001)。」於是,華萊士利用這種方法,以他所蒐集的鳥類資訊,在印尼數以萬計的島嶼中,畫出了一條線來,稱作「華萊士線」,線東的區域以亞洲大陸鳥類群相超過50%,稱作東洋區(Oriental),而線西則是大洋洲鳥類群相超過50%,稱作澳大拉西亞區(Australian)。

上圖就是華萊士線的位置。華萊士線後來又有其他學者加以補充或提出不同的看法,主要有韋伯線(Weber’s Line)、里德克線(Lydekker’s Line)和赫胥黎線(Huxley’s Line)。韋伯線可說是增補華萊士線的不足,將原本只採計鳥類的線重新計算為採計所有動物,大致在華萊士線的東側;里德克線則畫在澳洲大陸棚外圍,此線與華萊士線之間稱作「華萊士區」,在冰河時期是兩側陸棚間的海峽地區,基本上可算是兩大動物群相間的過渡地帶;赫胥黎線則是將華萊士線往北延伸至菲律賓一帶,又被稱作「新華萊士線」。不管哪條線,他們都代表著兩大動物群相的分界,而此一分界有意義的原因則在於,華萊士線大約標示著冰河時期海水面下降後,陸地分布的外圍,也就是說,華萊士線以東,在冰河時期仍是海洋,因此大陸動物無法渡海,與亞洲大陸的物種歧異度自然就相當的高。![]()
二十世紀初期,日本生物學家大島正滿提出了華萊士線往北延伸的可能性,並透過了日本探險家鹿野忠雄的實地調查發現,在蘭嶼的「球背象鼻蟲」與「蘭嶼光澤蝸牛」有相當程度的菲律賓生物相存在,而與台灣本島的生物相差異較大,繼續研究其他物種之後,提出華萊士線北延應該畫在台灣本島和蘭嶼、綠島之間。近代的生物學家研究蘭嶼、綠島的植物之後,也更加印證了這樣的想法。當然近代也有檢討,在島嶼生物地理學中,不同的物種有不同的擴散能力,這條線是否能這樣截然的劃分?不過,不可否認的,華萊士線對於後來的生物地理學研究,提供了相當重要的線索與指引,因此華萊士也被稱為「生物地理學之父」!
如果將來有機會,到蘭嶼、綠島去看看,別忘了仔細觀察一下動植物,尋找一下你所認識的華萊士線兩旁,到底有什麼不一樣!
侵蝕輪迴學說
(http://geog-education.blogspot.tw/2010/11/blog-post.html#more/游牧笛)
侵蝕輪迴學說 (Cycle of Erosion)


華倫亭海岸分類
(http://geog-education.blogspot.tw/2010/11/blog-post_1844.html#more/游牧笛)
在開始敘述華倫亭的海岸分類之前,可以先釐清一下,分類的目的到底是什麼?基本上將海岸地形做分類,不外乎下列幾個原因:第一,掌握海岸成因;第二,簡單了解海岸型態;第三,了解海岸發展。基本上,不論是為了商業交通目的(如建設港口)、海岸保安目的或者學術目的,海岸分類均十分重要。也正因為海岸分類十分重要,因此有許多人提出了不同的海岸分類,如沙岸、岩岸的分類,或者以成因而分成谷灣式(溺谷)海岸、珊瑚礁海岸……等。
華倫亭(H. Valentin),為了能夠解釋在地方的侵蝕、堆積作用與大環境的沉水、離水作用,因此提出了一套特別的海岸分類。所謂侵蝕、堆積指的是海浪、潮汐與洋流影響海濱地區,若海水作用力強,則以侵蝕作用為主,海作用弱則以堆積作用為主;沉水跟離水則是指整個大環境的海水面相對變化,如陸地相對抬升或海平面相對下降,則稱之為離水,如陸地相對下降或海平面相對升高,則稱作沉水。堆積與離水將會造成海岸線向海洋的方向移動,從人的角度看,就像是海岸線往前(往海的方向)前進,因此稱為「進夷」;反之,侵蝕與沉水將造成海岸線向陸地退縮,稱為「退夷」。
綜合了侵蝕、堆積、沉水、離水四大作用,華倫亭提出了一套他自己的海岸分類法,並可簡單繪製成下圖:

崩壞作用與崩壞地形
關於崩壞作用這篇文章講的精闢,值得一讀!
(http://geog-education.blogspot.tw/2010/11/blog-post_15.html/游牧笛)
這邊要淺談中學地理課會上到的幾種主要地形作用中的「崩壞作用」,以及這些作用所形成的地形。地形百百種,為什麼要先挑崩壞地形呢?有幾個主要原因:第一,先前在雪美老師的計畫中,曾經執行過原住民國中生對於坡地災害(崩壞地形所造成的災害)識覺的研究,發現學生對於崩壞作用的定義其實是相當模糊的;第二,當我進入高中實習之後,我也負責上到了崩壞地形,但很明顯在後來的驗收中,學生們似乎學的不是很好;第三,近年來崩壞地形所造成的災害,見報率與被研究的機率高了許多,提升這方面的認知確實有一定的必要,以免隨著部分記者以訛傳訛。
在一般狀況下,崩壞作用分為「崩塌」、「地滑」、「土石流」三種作用,而三民版的課本則在地滑中,將緩滑分出,稱作「潛移」,因此我們就暫且用這四種作用,來當作介紹的對象吧。
要弄清楚這幾個地形作用,要先搞清楚崩壞作用受到那些地形要素的影響。在戴維斯的地形理論中,構造、營力與時間三大要素是必須被討論的,所以我們先一項一項來釐清。
在構造上,崩塌一般而言發生在地質比較破碎的地點,例如斷層經過或者外營力侵蝕旺盛的位置;地滑則依其性質有所不同,坡面型地滑通常位於順向坡(尤其是坡腳被破壞處)的位置,而弧形地滑則通常發生在地質較為鬆軟處;土石流一般而言發生在岩屑較多的地方;潛移則好發於地面表層較鬆軟處。
在營力上,崩塌、地滑是以重力作用為主,若遇到大雨的潤滑或地震等導致震動,其發生的界檻值會降低,相對發生的機率就會增加;土石流主要以流水營力為主,配合上重力使得土石與水的混合能夠從河道、溝谷中流動;潛移也是以重力作用為主,受到水的潤滑等不明顯。
在時間上,崩塌、地滑都屬於快速發生,其中又以純重力作用所造成的崩塌速度最快,地滑居次,但也有快速如國道三號地滑一般的案例;土石流的速度差異大,其流速受到水量、坡度與谷床型態的影響,有慢到對生命安全無礙的,也有快如火車般可以直接沖進住家的;潛移是所有崩壞作用中速度最慢的,一般用肉眼無法觀察出來,多仰賴精密儀器,或者潛移發生處電線杆、墓碑的歪斜,或樹木基部的傾斜作為判斷指標。
受到不同的地形要素影響,不同的崩壞作用也會產生不同的崩壞地形。
崩塌通常是快速而近乎垂直的下落,崩落物堆積在坡腳或崖角處,常形成圓錐狀的「崩積錐」地形;坡面型地滑通常發生後,原地會出現岩層層面,地表平坦,弧形地滑則容易在發生位置上部出現小階梯狀的地形,下部則出現因重力擠壓而堆高的土丘;土石流在形態上,源頭的發生部經常為崩塌作用所產生的漏斗狀凹槽,接著順著溝谷流出,常造成河谷中有大量的「埋積物」,至平緩開闊的位置則類似沖積扇般,將沖積物展開而以扇狀平鋪於地表,稱作「土石扇」,土石扇與沖積扇的差別在於,沖積扇從扇頂到扇端顆粒明顯由大到小,而土石扇則是由小到大,但粒級分布並不顯著。
以下用幾張照片,把上述的概念解釋一下:


地滑速度快,大規模的地滑常造成地景的巨變。在台灣比較有名的例子,不外乎像是雲林草嶺地區因地震而滑落的事件,並且形成堰塞湖新草嶺潭,另一個有名的例子則是前陣子國道三號的地滑,造成了人員的傷亡;小林村的滅村事件,也可以算是個大規模的地滑。上述這兩種屬於坡面型的地滑,地滑發生的位置可以看到平滑的岩層層面。另一種地滑位於較鬆軟的地層(尤其可能發生在凸坡,也就是邊坡上部較緩,下部較陡處),例如苗栗的火炎山,該處岩層屬於頭嵙山層火炎山相,膠結程度較差,受到重力影響而在內部形成一個內凹的滑動面,在滑動區域上方可見到小階狀的景觀以及橫向裂隙,滑動區的下方則會有地表突起的現象。





